 המחקר שלנו מתמקד בהבנת המנגנונים המולקולריים שבאמצעותם מוטציות ברודופסין ובגנים המסדירים את סחר בה מובילות למחלות רשתית. רודופסין הוא מרכיב עיקרי של הממברנות החשות באור של תאי קולטני פוט של קרן רשתית, שם הוא מתפקד כ- GPCR לחישת אור היוזמת מפל פוטו-התמרה. רודופסין הוא גם וסת מרכזי לבריאות פוטורקולטורים וכפוף ליותר מ -150 מוטציות שונות הגורמות לרטיניטיס פיגמנטוזה. מוטציות המשפיעות על C-terminus של רודופסין גורמות לצורות חמורות של רטיניטיס פיגמנטוזה אוטוזומלית דומיננטית (ADRP). במהלך המחקר שלנו, הגדרנו את מוטיב ה- VxPx של מסוף ה- rhodopsin C כאות מיקוד ציליארי שמור (CTS). זיהינו את המרכיבים של מכלול המיקוד הציליארי שמזהה אות זה להסדרת סחר ברודופסין וחידוש קרום פוטורצפטר של מוט רשתית.
המחקר שלנו מתמקד בהבנת המנגנונים המולקולריים שבאמצעותם מוטציות ברודופסין ובגנים המסדירים את סחר בה מובילות למחלות רשתית. רודופסין הוא מרכיב עיקרי של הממברנות החשות באור של תאי קולטני פוט של קרן רשתית, שם הוא מתפקד כ- GPCR לחישת אור היוזמת מפל פוטו-התמרה. רודופסין הוא גם וסת מרכזי לבריאות פוטורקולטורים וכפוף ליותר מ -150 מוטציות שונות הגורמות לרטיניטיס פיגמנטוזה. מוטציות המשפיעות על C-terminus של רודופסין גורמות לצורות חמורות של רטיניטיס פיגמנטוזה אוטוזומלית דומיננטית (ADRP). במהלך המחקר שלנו, הגדרנו את מוטיב ה- VxPx של מסוף ה- rhodopsin C כאות מיקוד ציליארי שמור (CTS). זיהינו את המרכיבים של מכלול המיקוד הציליארי שמזהה אות זה להסדרת סחר ברודופסין וחידוש קרום פוטורצפטר של מוט רשתית.
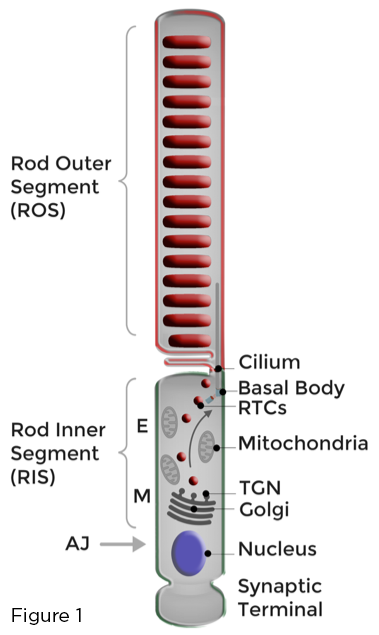 הבסיס לתפקוד האופטימלי של פוטורצפטורים של מוטים הוא המיון הקפדני של הממברנות הרגישות העמוסות לרודופסין לציליום העיקרי שהשתנה באופן ייחודי היוצר את המקטע החיצוני של המוט (ROS). הומאוסטזיס של קולטן פוט מקיים על ידי חידוש מתמשך של ממברנות ROS שנפגעו באור ורצף יעיל של חלבוני ROS ושומנים המעורבים בפוטו-הולכה, הרחק מאלה העוסקים בפונקציות סלולריות מגוונות המתקיימות בגוף התא הסמוך ושמו קטע הפנימי של המוט (RIS), או במסוף הסינפטי של המוט. גולגי ואברונים ביו -סינתטיים אחרים ממוקמים ב- RIS, באזור המיואיד (M). סחר בממברנה מקוטבת במוטות ברשתית כרוך בסינתזה, מיון והובלה באמצעות ה- RIS של כמות אדירה של נושאי הובלת רודופסין (RTCs) המכוון על ידי גולגי. RTCs חוצים את אזור האליפסואיד (E) המלא במיטוכונדריה ומתמזגים עם קרום הפלזמה של RIS בסמיכות הציליום. ממברנות מסונתזות חדשות מועברות לאחר מכן ל- ROS (איור 1).
הבסיס לתפקוד האופטימלי של פוטורצפטורים של מוטים הוא המיון הקפדני של הממברנות הרגישות העמוסות לרודופסין לציליום העיקרי שהשתנה באופן ייחודי היוצר את המקטע החיצוני של המוט (ROS). הומאוסטזיס של קולטן פוט מקיים על ידי חידוש מתמשך של ממברנות ROS שנפגעו באור ורצף יעיל של חלבוני ROS ושומנים המעורבים בפוטו-הולכה, הרחק מאלה העוסקים בפונקציות סלולריות מגוונות המתקיימות בגוף התא הסמוך ושמו קטע הפנימי של המוט (RIS), או במסוף הסינפטי של המוט. גולגי ואברונים ביו -סינתטיים אחרים ממוקמים ב- RIS, באזור המיואיד (M). סחר בממברנה מקוטבת במוטות ברשתית כרוך בסינתזה, מיון והובלה באמצעות ה- RIS של כמות אדירה של נושאי הובלת רודופסין (RTCs) המכוון על ידי גולגי. RTCs חוצים את אזור האליפסואיד (E) המלא במיטוכונדריה ומתמזגים עם קרום הפלזמה של RIS בסמיכות הציליום. ממברנות מסונתזות חדשות מועברות לאחר מכן ל- ROS (איור 1).
המיון ל- RTCs מוסדר על ידי GTPases הקטנים של משפחות Rab ו- Arf, הממלאים תפקיד מרכזי בארגון סחר בממברנות תוך תאי וכן משלוח ממברנה לציריות הראשוניות. חלבונים המעורבים בהיווצרות ותחזוקה של ציליליה מקודדים על ידי כ -25% מהגנים התורשתיים של מחלת הרשתית, כאשר מוטציות גורמות להתנוונות הרשתית, כליות ציסטיות, השמנת יתר ופגמים בצינור העצבי במגוון רחב של הפרעות גנטיות, הידועות ביחד בשם ciliopathies.
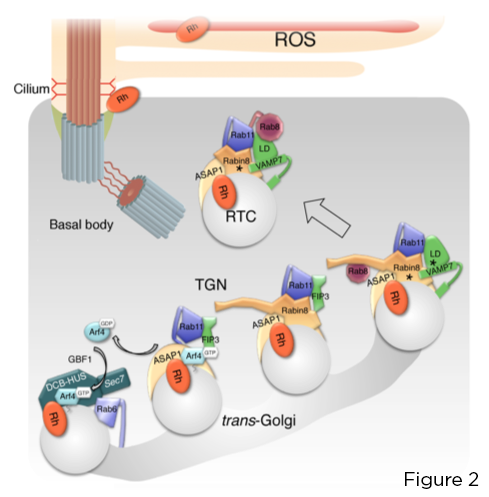 ה- GTPase הקטן Arf4 מזהה וקושר ישירות את ה- C-terminale VxPx C-rhodopsin. Arf4, המופעל על ידי קולטן הצילום Golgi על ידי גורם חילופי נוקלאוטיד Arf גואנין (GEF) GBF1, יוזם הרכבה שלבים של קשרי המיקוד שבמרכזו החלבון הפועל Arf GTPase (GAP) ASAP1, המתווך הידרוליזה של GTP ב- Arf4, ו- Rab11a -מתחם אפקטור כפול FIP3-Rabin8. קומפלקס זה שולט בהרכבה של מודול המיקוד Rab11a-Rabin8-Rab8 שמור מאוד שמגייס ישירות את ה- R-SNARE VAMP7 אל RTCs כדי להסדיר את ההתמזגות שלהם בבסיס הציליארי, באמצעות זיווג VAMP7 עם סינטקסין 3 ו- SNAP-25 ממברנת הפלזמה המזוהה. (איור 2).
ה- GTPase הקטן Arf4 מזהה וקושר ישירות את ה- C-terminale VxPx C-rhodopsin. Arf4, המופעל על ידי קולטן הצילום Golgi על ידי גורם חילופי נוקלאוטיד Arf גואנין (GEF) GBF1, יוזם הרכבה שלבים של קשרי המיקוד שבמרכזו החלבון הפועל Arf GTPase (GAP) ASAP1, המתווך הידרוליזה של GTP ב- Arf4, ו- Rab11a -מתחם אפקטור כפול FIP3-Rabin8. קומפלקס זה שולט בהרכבה של מודול המיקוד Rab11a-Rabin8-Rab8 שמור מאוד שמגייס ישירות את ה- R-SNARE VAMP7 אל RTCs כדי להסדיר את ההתמזגות שלהם בבסיס הציליארי, באמצעות זיווג VAMP7 עם סינטקסין 3 ו- SNAP-25 ממברנת הפלזמה המזוהה. (איור 2).
בבסיס המפל הצלילי Rab11a-Rabin8-Rab8 נמצא ה- Rab8 GEF Rabin8, חלבון פיגום רב תכליתי המתקשר עם חלבונים ציליאריים נבחרים, כגון מתחם הסחר ב- TRAPPII ו- BBSome, דבר המצביע על תפקיד מרכזי במסלולים ריצוניים של קולטנים חושיים. תפקודו מושפע ממוטציות ב- NDR2 kinase (STK38L), המקודדות על ידי הגן הניוון המוקדם של הרשתית (erd) הכלב המתאים ל ciliopathy האנושי לבור אמורוזיס (LCA). המחקר הנוכחי שלנו מראה ש- GFP-Rabin8 אנושי מתבטא בטרנסגניים X. laevis מתמקדת עם Rabin8 אנדוגני ורודופסין בגולגי וב- RTCs, וסוללת את הדרך למחקרים עתידיים על תפקידו של Rabin8 בהתקדמות הממברנה לאורך המסלול הצילירי, שעלול להפריע במחלות ניווניות ברשתית.
יחד, מחקרינו גילו כי מיקוד ממברנות ל- ROS הוא צורה שמורה של מיקוד ציליארי. מוטיב ה- VxPx קיים בחלבוני קרום רירית אחרים. קומפלקס המיקוד המבוסס על Arf4 ממקד קולטנים חושיים לציריות ראשונות באמצעות רשתות תפקודיות מורכבות של GTPases קטנים והרגולטורים שלהן הרגישים להפליא למוטציות הגורמות להתנוונות רשתית וסיליופתיות.
Deretic D., Lorentzen E ו- Fresquez T. (2019). החלקים הפנימיים של מכלול מיקוד הממברנה הסילירית המבוססת על Arf4. GTPases קטנים. GTPases קטנים. 2019 9 במאי: 1-12. doi: 10.1080/21541248.2019.1616355. [Epub לפני ההדפסה]
Kandachar V, Tam BM, Moritz OL ו- Deretic D. (2018) רשת אינטראקציה בין SNARE VAMP7 ו- Rab-GTPases בתוך קומפלקס מיקוד קרום ריאלי. J Cell Sci. (2018) J Cell Sci. 2018 10 בדצמבר; 131 (24). pii: jcs222034. doi: 10.1242/jcs.222034.
וואנג ג ', פרסקז ט. קנדצ'אר ו. ודרטיק ד (2017) ה- Arf GEF GBF1 ו- Arf4 מסתנכרים עם מטען הקולטן החושי, רודופסין, כדי להסדיר את סחר בקרום הרירית. J Cell Sci. J Cell Sci. 2017 1 בדצמבר; 130 (23): 3975-3987. doi: 10.1242/jcs.205492. Epub 2017 12 באוקטובר.
Vetter M, Wang J, Lorentzen E ו- Deretic D. (2015) טופוגרפיה חדשה של רשת האינטראקציה של Rab11-effektor בתוך קומפלקס ממוקד ממברנה רירית. GTPases קטנים. 2015 2 באוקטובר; 6 (4): 165-73. doi: 10.1080/21541248.2015.1091539. Epub 2015 23 בספטמבר.
וואנג ג 'ודרטיק ד' (2015) FIP11 מפעיל Arf ו- Rab3 פועל באופן סינרגיסטי עם ה- Arf GAP ASAP1 כדי לכוון את רבין 8 במיקוד קולטן רילי. J Cell Sci. J Cell Sci jcs.162925; מאמר מקוון מראש 11 בפברואר 2015, doi: 10.1242/jcs.162925 (2015).
וואנג ג'יי ודרטיק ד (2014). מתחמים מולקולריים המכוונים את הובלת הרודופסין לציריות הראשוניות. Prog Retin Eye Res. 2014 בינואר; 38: 1-19. doi: 10.1016/j.preteyeres.2013.08.004. Epub 2013 14 באוקטובר
וואנג ג'יי, מוריטה Y, Mazelova J ו- Deretic D. (2012). ה- Arf GAP ASAP1 מספק פלטפורמה לוויסות מיקוד קולטן ריאלי בתיווך Arf4 ו- Rab11-Rab8. EMBO י 31, 4057-4071.
Mazelova J, Ransom N, Astuto-Gribble L, Wilson MC ו- Deretic D. (2009) זיווגי Syntaxin 3 ו- SNAP-25, המווסתים על ידי חומצה דוקוזהקסאנואית אומגה 3 (DHA), שולטת במתן רודופסין לביוגנזה של ציליליה- אברונים תחושתיים נגזרים, הקטעים החיצוניים של המוט. J. Cell Sci. 122, 2003-2013.
Mazelova J, Astuto-Gribble L, Inoue H, Tam BM, Schonteich E, Prekeris R, Moritz OL, Randazzo PA ו- Deretic D. (2009) מוטיב הכוונה Ciliary VxPx מכוון הרכבה של מודול סחר דרך Arf4. EMBO J. 28, 183-192.
Deretic D, Williams AH, Ransom N, Morel V, Hargrave PA ו- Arendt A. (2005) Rhodopsin C-terminus, אתר המוטציות הגורמות למחלות רשתית, מסדיר את הסחר על ידי קישור ל- ARF4. מעבד נטל. אקאד. Sci. ארה"ב. 102: 3301-3306.